
A transcrição consiste na síntese de uma molécula de RNA a partir de um molde de DNA. Na molécula de DNA, bifilamentar, um dos filamentos contém a informação, sendo chamado fita codificadora. O outro filamento, que é utilizado como modelo para a síntese, é denominado fita molde.
A síntese de RNA é realizada por uma enzima, a RNA polimerase. Para que ocorra a transcrição é necessario que a RNA polimerase reconheça uma região específica do gene, que delimita o ponto de início da informação. Esta região é chamada de promotor. Nos procariotos, o promotor apresenta duas sequências importantes para o reconhecimento, localizadas a -10 e -35 nucleotídeos do ponto de início da transcrição (denominado por convenção como +1). Nos eucariontes a estrutura do promotor é mais complexa, com um número maior de sequências de reconhecimento (que incluem sequencias a -25; -75; -90 e -100 nucleotídeos em um promotor basal).
A RNA polimerase
Em procariotos, há apenas um tipo de RNA polimerase, que pode se apresentar em duas formas: apoenzima e holoenzima. A diferença entre estas formas da RNA pol é a presença do fator sigma, uma das subunidades peptídicas que compõe a RNA pol. A apoenzima não possui fator sigma, ao passo que a holoenzima possui.

A presença do fator sigma habilita a enzima a reconhecer a região promotora dos genes a serem transcritos e a se ligar ao DNA. Entretanto, este mesmo fator sigma dificulta o exercício da atividade polimerásica pela enzima. Desta forma, após reconhecer o promotor e se ligar ao DNA, a holoenzima perde o fator sigma, convertendo-se em apoenzima. A apoenzima, por sua vez, possui elevada processividade na síntese, ao passo que não tem habilidade para reconhecer o promotor dos genes. Desta maneira, a holoenzima reconhece os genes a serem transcritos e se liga ao DNA e a apoenzima faz a síntese do RNA.

Em eucariontes observamos 3 tipos de RNA polimerase: a RNA pol I, responsável pela síntese de rRNA (RNA ribossomal); a RNA pol II, responsável pela transcrição do mRNA (RNA mensageiro) e a RNA pol III, responsável pela sintese de tRNA (RNA transportador) e outros de pequena extensão. estas polimerases não reconhecem diretamente o promotor, necessitando do auxílio de proteínas que possuem domínios de ligação ao DNA e são chamadas fatores transcricionais. Ao se ligarem ao promotor, os fatores de trancrição promovem a formação da geometria de ligação requerida pela RNA polimerase para se associar ao DNA.

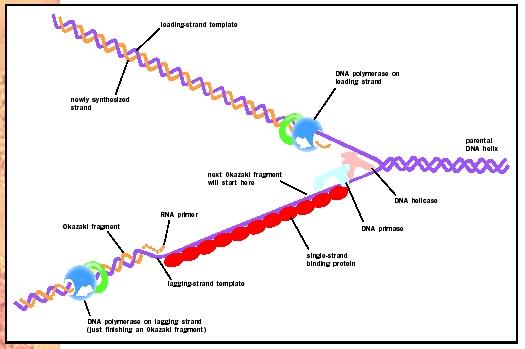
Após reconhecer a região promotora, a RNA polimerase encadeia os nucleotídeos de forma semelhante à DNA polimerase, ou seja, apresenta atividade polimerásica 5´- 3´. Lembro aqui que o RNA não possui timina, sendo a uracila encadeada em sua substituição. Esta etapa é conhecida como alongamento de cadeia.
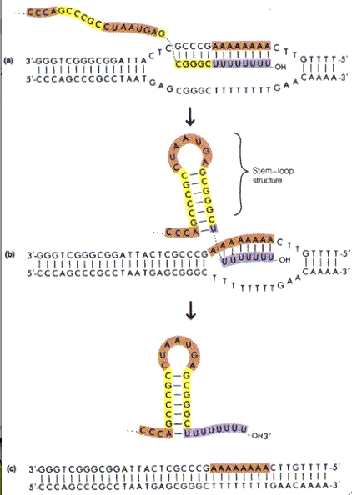
No fim da trancrição, etapa denominada terminação, observamos diferenças entre procariontes e eucariontes. nas bactérias há duas estratégias de terminação, requerendo ou não a participação de uma proteína chamada rho. Na terminação independente de rho, também denominada terminação intrinseca, há uma sequência rica em pares A/U logo após a região que delimita o fim da transcrição, permitindo que a RNA pol se libere do DNA naturalmente. Já na terminação dependente de rho, a presença de uma região rica em pares C/G impede que a polimerase se libere naturalmente, havendo necessidade da ação da proteína rho, que libera a enzima.
Nos eucariontes, o termino da transcrição ocorre de forma indeterminada, havendo liberação da RNA polimerase em uma região variável após a sequência de poliadenilação.
Nos eucariontes, o termino da transcrição ocorre de forma indeterminada, havendo liberação da RNA polimerase em uma região variável após a sequência de poliadenilação.
Apesar de também poder haver correção de erros na transcrição, as RNA polimerases não apresentam a mesma eficiência no processo que as da DNAs polimerases (que possuem atividade exonucleásica). A correção no processo de transcrição pode ser feita atraves de i) correção cinética (ou pirofosfólise) e ii) correção nucleolítica e a taxa de erro fica em torno de 10-4 a 10-5.
pirofosfólise

correção nucleolítica

Tipos de RNA
Os principais tipos de RNA observados nas células procariontes e eucariontes são:
1) RNA mensageiro (mRNA) - responsável pelo carreamento da informação contida na molécula de RNA


3) RNA transportador (tRNA) - responsável pelo carreamento de aminoácidos através do citoplasma para o sítio de tradução
Estes RNAs atuam cooperativamente para que ocorra a síntese de proteínas

Nas células eucarióticas, temos ainda:
4) RNA heterogêneo nuclear (hnRNA) –transcritos primários que possuem regiões contendo informações para codificação (EXONS) e regiões que não possuem tais informações (INTRONS) que irão gerar, após o processamento, o mRNA.
5) RNA pequeno nuclear (snRNA – small nuclear RNA) – RNAs de pequeno tamanho (cerca de 70 nucleotídeos), ricos em uracila (recebem genericamente a designação de Un), que participam da remoção dos introns do hnRNA. Ocorrem na forma de ribonucleoproteínas (SNRPs ou SNURPs).
6) RNA pequeno citoplasmático (scRNA – small cytoplasmic RNA) – pequenas moléculas de RNA, geralmente com cerca de 130 nucleotídeos, que ocorrem no citoplasma e em associação ao retículo endoplasmático e potencialmente associadas com proteínas envolvidas em transporte de mRNA (?) e seleção de proteínas. Ocorrem na forma riboproteínas (SCYRPs).
Achou que havia acabado? Se enganou:
7) MicroRNA (miRNA) – RNAs de cadeia curta (com 20 a 30 nucleotídeos) que se associam a um complexo proteico (RISC – RNA induced silencing complex) para reprimir a expressão de genes alvo, através da formação de duplexes na região 3´não traduzida do mensageiro.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}