REPLICAÇÃO DO DNA
O
DNA apresenta características estruturais únicas que o tornam adequado
para ser a molécula da hereditariedade, como o fato de ser bifilamentar e
a interação específica entre as bases nitrogenadas, através das pontes
de hidrogênio estabelecidas.
O modelo estrutural proposto por Watson e
Crick para a molécula de DNA sugeria que a replicação do DNA fosse realizada de uma forma
tal que cada filamento, individualmente, fosse utilizado como modelo
para síntese de seu filamento complementar. A demonstração experimental
foi feita por Meselson e Stahl (1958), em experimentos utilizando incorporação de nitrogênio com densidade diferenciada. Este trabalho foi muito importante para consolidar a aceitação
sobre a proposição molecular de Watson e Crick. No estudo, os
pesquisadores cultivaram células bacterianas por várias gerações em meio
de cultivo contendo apenas N15 disponível para síntese de biomoléculas.
Ao transferirem as bactérias para meio contendo N14, a formação de
moléculas híbridas de DNA, contendo um filamento com N14 e um filamento
com N15 foi a base para a determinação do modelo SEMICONSERVATIVO de duplicação.
Desta forma, uma molécula de DNA SEMPRE apresenta um filamento velho (oriundo da molécula que a originou) e um filamento novo (recém sintetizado).
{kind=link}
Desta forma, uma molécula de DNA SEMPRE apresenta um filamento velho (oriundo da molécula que a originou) e um filamento novo (recém sintetizado).

A enzima responsável pela síntese de DNA é a DNA polimerase. Em procariotos, encontramos 3 tipos de DNA polimerase: DNA pol I; DNA pol II e DNA pol III. Através de experimentação com mutantes bacterianos, sabe-se que DNA pol I tem papel representativo na correção de erros e que DNA pol III é a principal enzima para síntese de DNA (recentemente, duas novas polimerases, DNA pol IV e DNA pol V, ambas associadas com reparo de lesões no DNA foram inseridas neste rol). Em eucariotos há um grande número de polimerases. Para replicação, são necessárias as polimerases alfa, delta e epsilon. A polimerase beta está associada a correção de erros e a polimerase gama é responsável pela replicação do DNA mitocondrial. Caracteristicamente as DNA polimerases apresentam atividade polimerásica 5´-3´ e atividade exonucleásica 3´-5´. Esta última assegura a correção de possíveis erros de pareamento durante a síntese de DNA. O sistema funciona pela competição dos sítios catalíticos: Como a afinidade do sítio polimerásico é maior que a do sítio exonucleásico, apenas quando o nucleotídeo alocado não realiza o pareamento adequado é possível removê-lo. Há, ainda, a atividade exonucleásica 5´-3´ exibida pela DNA polimerase I, que permite a remoção dos iniciadores e auxilia no reparo a erros identificados após a replicação.
A descoberta dos mecanismos de síntese biológica dos ácidos nucleicos rendeu o prêmio Nobel de 1959 a Severo Ochoa e Arthur Kornberg. Para que ocorra a replicação é necessária a ação de uma série de enzimas
e proteínas que prepararão a molécula de DNA para a ação da polimerase.
A região na qual a replicação se inicia é denominada origem de
replicação (ori). Em procariontes, esta região ocupa um segmento
de cerca de 220pb, sendo constituída por sequências conservadas
dispostas de maneira específica na molécula de DNA. Na ori observamos a
ocorrência de 3 repetições de uma sequência conservada de 13
nucleotídeos (GATCTNTTNTTTT) seguida de quatro repetições de uma
sequência de 9 nucleotídeos (TTATNCANA). Nesta região é formada a bolha
de replicação através da ação do aparato enzimático da replicação.
{kind=link}

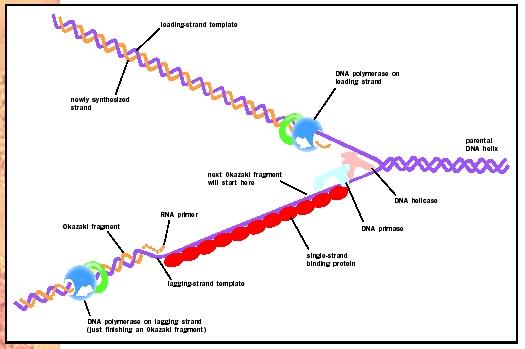
A girase promove a torção negativa necessária para que a molécula de DNA possa ser duplicada. Já a helicase tem o papel de romper as pontes de hidrogênio entre as bases nitrogenadas. As proteínas SSB estabilizam os filamentos de DNA separados e a primase realiza a síntese do iniciador ou primer. Assim forma-se um complexo replicativo que permite a ação da DNA polimerase e a forquilha de replicação avança para que a replicação tome curso. Como os filamentos do DNA são antiparalelos, enquanto uma das fitas é sintetizada de forma contínua, a outra fita é sintetizada em pedaços, denominados fragmentos de Okazaki. Assim, a replicação, além de semiconservativa, é SEMIDESCONTÍNUA. A ligase tem papel de unir os segmentos sintetizados para que a replicação possa ser concluída. é importante lembrar que, atraves da atividade exonucleásica 5´-3´, o iniciador é removido, sendo substituído por um segmento de DNA.
{kind=link}

Nas
células eucariontes, como o DNA é linear, uma das extremidades da fita
descontínua terminal não é completamente duplicada pelo aparato de
replicação do replissoma. Nesta região age uma enzima ribonucleoproteica
especial, descoberta na década de 80, denominada telomerase. A
descoberta foi agraciada com o prêmio Nobel em 2009!
{kind=link}

Seus principais componentes são codificados pelos genes TLC1 (molde de RNA) e EST2 (porção catalítica). Esta enzima utiliza seu próprio constituinte de RNA para servir de molde à duplicação das sequências repetitivas TTAGGG/TTGGGG que compõe o telômero.

A
atividade telomerase é observada em todas as células em divisão, sendo
diminuída progressivamente em células com maior diferenciação.
Oi Marcelo, não entendi muito bem sobre o fragmento Klenow no papel da replicação. Poderia me explicar na próxima aula ? Obrigada !
ResponderExcluir